1 材料与方法
1.1 研究区概况
研 究 区 位 于 新 疆 塔 里 木 盆 地 西 北 边 缘(81°17'56.52″E,40°32'36.90″N)。研究区气候炎热干燥,多年平均降雨量仅 50 mm 左右,潜在蒸发量可达 1 900 mm,年均气温 10.8 ℃,年均日照时数为2 900 h,是典型的温带荒漠气候。
1.2 试验设计
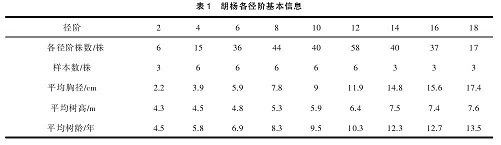
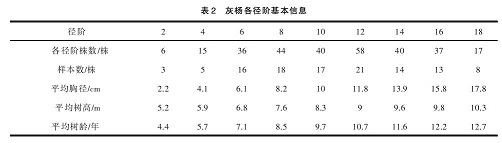
本研究以不同径阶的胡杨、灰杨代表其不同的发育阶段。径阶划分以及标准木确定方法为:以胸径相差2 cm为标准进行规划,起始径阶为2 cm。将研究区内 355株胡杨划分为 2径阶、4径阶、6径阶、8径阶、10径阶、12径阶、14径阶、16径阶、18径阶(9个径阶代表了胡杨9个发育阶段)。研究区内共有灰杨301株,划分方法与胡杨相同,在胡杨、灰杨2~18 cm径阶分别选取3株作为重复,胡杨、灰杨标准木各27株见表1、表2。
各径阶胸径和树龄之间的关系如下公式:
A=13.679/ (1+3.347 6 ×exp(-0.209 9 D))其中: A是树龄,D是胸径。
1.3 采样方法
胡杨、灰杨叶片发育成熟时期,将植株冠高(树高-枝下高)5 等分,定义 5 等分后由树冠基部向顶部方向依次为树冠 1~5 层。在树冠 5 个层次的中央位置,按东、南、西、北方位各随机采取 1个当年生枝条,每株标准木共采集 20个枝条。选取每个枝条基部向顶端方向的第三个叶片为样叶,每一个层次 4 个叶片,每株标准木共计 20个叶片,用于异形叶形态测定和制作组织切片。
1.4 叶片形态结构指标测定方法
异形叶形态测定方法:将当年生枝条上的叶片按节位带叶柄取下,以 MRS-9600TFU2 扫描仪对叶进行扫描,用万深 LA-S 型植物图像分析软件进行叶片长、叶片宽、叶面积、叶柄长、叶周长、叶片厚度、叶形指数(叶片长/宽比值)的测量。
异形叶解剖结构测定方法:取当年生枝条上第 3节位的叶,切取异形叶最宽处材料以 FAA 固定液保存。采用常规石蜡制片方法制作组织切片,切片厚度 8 μm、番红固绿双重染色,中性树脂封片。在 Lei⁃ca 显微镜下观察测量异形叶最宽处横切面表皮组织(表皮细胞数、表皮细胞长度和宽度)、栅栏组织(栅栏组织厚度、栅栏组织细胞长度和宽度)及海绵组织(海绵组织厚度)结构参数。计算栅栏组织厚度与海绵组织厚度的比值(栅/海比值)。每叶片观测 5 个视野,每视野观测 20 个值,取 5 个视野叶片结构参数的平均值为每叶片结构参数值。
1.5 数据处理
各发育阶段异形叶形态、解剖结构指标参数为各径阶 3株标准木异形叶形态、解剖结构指标参数值的平均值。
采用SPSS 17.0软件数据处理系统对胡杨、灰杨9个径阶异形叶 7 个形态性状和 9 个解剖结构性状进行主成分分析,确定不同径阶异形叶形态结构差异的主要性状。常用描述统计量的计算和数据转换使用 Microsoft Office Excel 2003 软件。变异系数(CV)、主成分分析均采用SPSS 17.0软件。
2 结果与分析
2.1 胡杨异形叶形态结构性状主成分分析
2.1.1 胡杨异形叶形态性状主成分分析
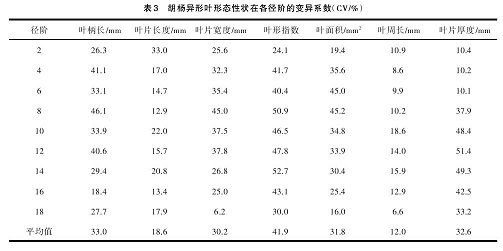
从表 3 看出,胡杨异形叶 7 个形态性状在各径阶的变异系数平均值,从大到小依次为:叶形指数、叶柄长度、叶片厚度、叶面积、叶片宽度、叶片长、叶周长,说明各径阶均是叶形指数变异最大(CV=41.9%),说明不同径阶的胡杨异形叶形态差异较大;其次是叶柄长度、叶片厚度、叶面积变异系数较大。叶片宽度变异系数大于长度变异系数,表明叶片宽度变化比长度变化更丰富,而叶片长宽的变化间接影响到叶面积的变化,从而导致叶面积的变异较大。
从各径阶 7个形态性状变异系数可以看出,胡杨异形叶形态性状变异系数从 2 径阶至 18 径阶呈现“抛物线”式的变化,说明 8~14 径阶是胡杨异形叶形态性状变异比较大的阶段,而 18 径阶叶形态性状的变异系数变化的幅度,是所有径阶中最小的,说明 18径阶胡杨异形叶形态性状趋于稳定。由此表明,胡杨异形叶形态性状的变异在不同径阶有所不同,异形叶形态性状变异具有阶段性的特点。
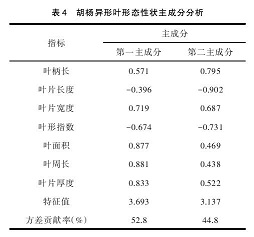
由表 4 可知,前两个主成分占总变异的 97.6%,其中第一主成分对总变异的贡献率为 52.8%,7 个形态性状对第一主成分影响较大的三个性状有叶周长、叶面积、叶片厚度;第二主成分对总变异的贡献率为 44.8% ,对第二主成分影响最大形态性状为叶片长度。总的看来,叶周长、叶面积、叶片厚度、叶片长度 4个性状影响异形叶形态变化,是造成胡杨不同径阶异形叶形态性状差异的主要内在因素。
2.1.2 胡杨异形叶解剖结构性状主成分分析
从表 5可以看出,胡杨异形叶解剖结构性状在各径阶的变异系数具有明显的差异,叶片解剖结构性状中 9个指标均有变异,其中变异系数最大的是栅栏组织细胞数和表皮细胞数。
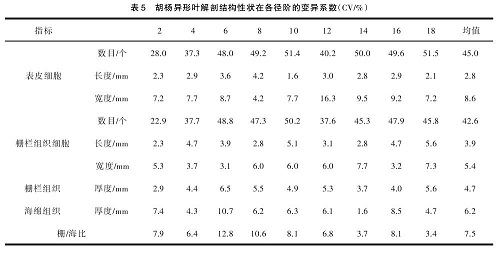
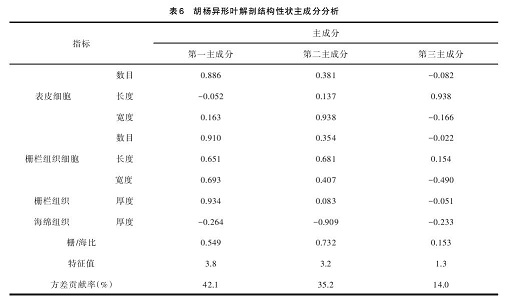
从表 6 可见,前 3 个主成分构成的信息量占总信息量的 91.3%。其中第一主成分对总变异的贡献率为 42.1%,对第一主成分影响最大的结构性状是叶片的栅栏组织厚度;第二主成分对总变异的贡献率为35.2%,对第二主成分影响最大的结构性状是海绵组织厚度;第三主成分占总变异的 14.0%,表皮细胞长度具有最大的影响。结果表明,表皮细胞长度、海绵组织厚度、栅栏组织厚度是造成不同发育阶段胡杨 9个径阶异形叶解剖结构差异的主要因素。
2.2 灰杨异形叶形态结构性状主成分分析
2.2.1 灰杨异形叶形态性状主成分分析
对灰杨异形叶的 7个形态性状进行测量统计,发现灰杨异形叶的 7 个形态性状在各径阶都存在不同程度的变异(表 7),变异系数平均值从大到小依次为: 叶柄长、叶面积、叶形指数、叶片宽度、叶片厚度、叶周长、叶片长度。变异系数最大的是叶柄长(CV=12.4%)说明不同径阶间异形叶叶柄长度变异较大;其次变异系数较大的是叶面积(CV=11.0%),叶片宽度变异系数大于叶片长度变异系数,当叶片长度变异最小时,叶片宽度变异可能就成为影响叶面积变异的主要因素。
从各径阶 7 个形态性状变异系数可以看出,12~14 径阶异形叶形态性状变异幅度最大的阶段,而8 径阶异形叶形态性状变异幅度最小。说明该阶段异形叶形态性状比较稳定。由此说明,灰杨异形叶形态性状的变异在不同径阶存在差异。
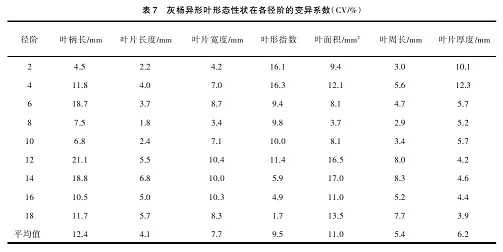
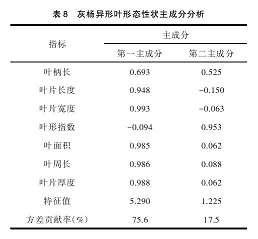
表 8 表明提取的主成分占总变异的 93.1%,其中第一主成分占总变异的 75.6%,影响较大的有:叶片宽度、叶片厚度、叶面积、叶周长;第二主成分对总变异的贡献率为 17.5% ,对其影响最大的性状为叶形指数。总的看来,叶周长、叶片厚度、叶面积、叶片宽度与叶形指数 5 个性状是造成灰杨不同径阶异形叶形态性状差异的主要因素。
2.2.2 灰杨异形叶解剖结构性状主成分分析
灰杨异形叶解剖结构性状在各径阶的变异系数如表 9,在各径阶中,异形叶解剖结构性状均是栅栏组织细胞数和表皮细胞数变异系数最大,说明各径阶异形叶解剖结构的变化主要体现在栅栏组织细胞数和表皮细胞数的变化上。
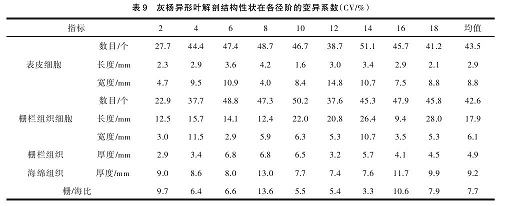
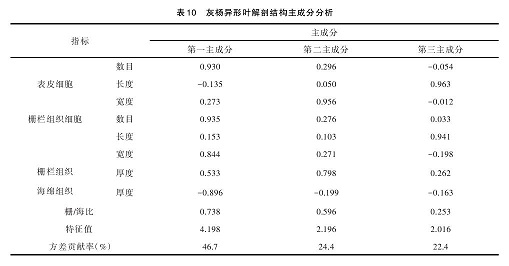
从表 10 可以得出,提取出的信息量占总信息量的 93.5%。 其 中 第 一 主 成 分 占 总 变 异 贡 献 率 的46.7%,对第一主成分影响最大是表皮细胞数、栅栏组织细胞数、海绵组织厚度;第二主成分对总变异的贡献率为 24.4%,对第二主成分影响最大的结构性状是表皮细胞宽度、栅栏组织厚度;第三主成分占总贡献率的 22.4%,对第三主成分影响最大的性状是表皮细胞长度、栅栏组织细胞长度。表皮细胞数、海绵组织厚度、栅栏组织厚度及栅栏组织细胞数是决定灰杨不同发育阶段异形叶结构差异的主要内在因素。
2.3 胡杨、灰杨异形叶形态结构性状与径阶相关性分析
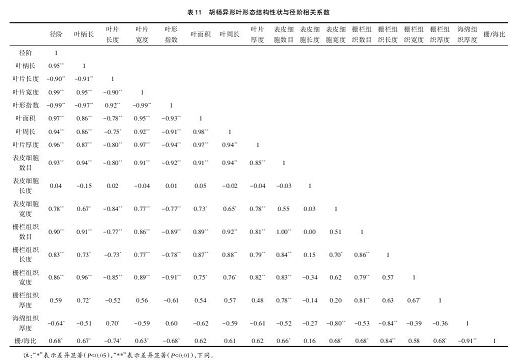
表 11 显示,胡杨叶柄长、叶片宽度、叶面积、叶周长、叶厚度、表皮细胞数目、表皮细胞宽度、栅栏组织数目、栅栏组织长度、栅栏组织宽度与径阶呈极显著正相关(P<0.01),栅海比与径阶呈显著正相关(P<0.05),而叶片长度、叶形指数与径阶呈极显著负相关,海绵组织厚与径阶呈显著负相关,结果表明胡杨异形叶形态结构性状与径阶有密切的相关性,即与胡杨不同发育阶段有关联。
从表 12可以看出灰杨叶柄长、表皮细胞数目、表皮细胞宽度、栅栏组织数目、栅栏组织宽度、栅栏组织厚度、栅海比与径阶呈极显著正相关(P<0.01),叶面积、叶周长与径阶呈显著正相关(P<0.05),而海绵组织厚度与径阶呈极显著负相关。表明叶柄长、叶面积、叶周长及在叶片最宽处横切面表皮细胞长度、表皮细胞宽度、栅栏组织长度、栅栏组织厚度、海绵组织厚度、栅海比随径阶的增加而增加。说明灰杨形态结构性状与径阶具有一定的关联性。
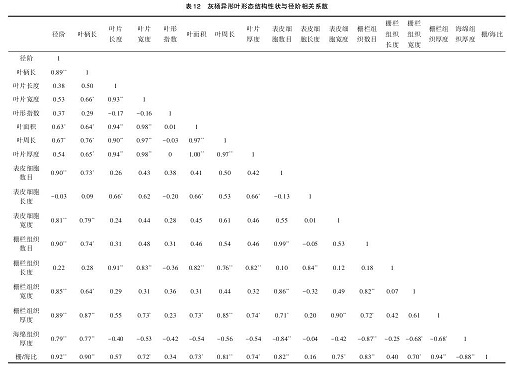
结合表 11、表 12 来看,胡杨、灰杨异形叶形态结构性状与径阶具有密切的相关性,随径阶的变化而变化,并且在胡杨、灰杨个体间的异形叶形态结构形状参数间也有极显著的正、负相关性。说明胡杨、灰杨个体间的异形叶形态结构形状指标也存在着相辅相成的协同变化关系。
原文引用
曲文蕊,韩晓莉,翟军团,李志军.不同发育阶段胡杨、灰杨异形叶形态结构变化特征[J].塔里木大学学报,2021,33(02):14-24.